|
Pasningsvejledning • Opdrćt • Genetik • Litteratur Vejledning i genetik. 2. udgaveaf Thomas GrochalskiForordI denne nye udgave gĺr jeg langt mere i detaljer med, hvordan tingene foregĺr, end jeg gjorde i den tidligere vejledning. Jeg har desuden tilfřjet flere ekstra interessante emner. Jeg gennemgĺr dog samtidig alt stof som i den tidligere udgave, men pĺ et lidt hřjere niveau.I teksten vil jeg henvise til opgaver man kan bruge pĺ at teste sig selv (Henvisninger ikke tilfřjet endnu). Opgaverne stĺr i nedenstĺende dokument, og svarene i det efterfřlgende dokument. Genetik.pdf Genetik_SVAR.pdf IndholdsfortegnelseDe emner som er interessante, men ikke strengt nřdvendige til kornsnogegenetik er markeret med rřdt (orange kan vćre nřdvendigt, men kun ved pied-sided). Punkterne i indholdsfortegnelsen fungerer som hyperlinks til de specifikke sektioner.
Introduktion til genetikLigesom vores arvemateriale er bestemt af hver vores specifikke DNA-kode er det samme tilfćldet for slanger. Langt střrstedelen af vores DNA-kode er fordelt pĺ kromosomer, hvilket er den type DNA, vi beskćftiger os med i morfavl. Der findes ogsĺ mitokondriel DNA (mtDNA), men det vil jeg ikke fokusere pĺ her.DNA'en er fordelt pĺ flere kromosomer. Der er to typer kromosomer - křnskromosomer (allosomer) og autosomer. Kornsnoge er en diploid organisme, hvilket betyder at den har to kopier af hvert autosom plus to křnskromosomer. Kornsnoge har i alt 36 kromosomer, hvilket betyder 17 par autosomer plus 2 křnskromosomer. De arver et sćt fra hver forćlder, dvs de modtager 17 autosomer og et křnskromosom fra hver forćlder. Et gen er en specifik sektion af DNA-strengen, som koder for et bestemt trćk - det kunne f. eks vćre melaninproduktion. Der kan sĺ vćre forskellige variationer af et gen. Hvis vi antage,r det er melaninproduktionen, vi kigger pĺ, kan det f. eks vćre albinisme eller vildtypen (normalen). Dette er allele former af det gen, dvs. allele former betyder reelt de forskellige variationer, der er af et specifikt gen. Den placering pĺ kromosomet, som genet er placeret, hedder et locus. Nĺr vi arbejder med morfer er det de allele former af specifikke gener som vi ser nćrmere pĺ. Nĺr vi har et kromosompar, betyder det ogsĺ at vi har to kopier af det samme gen. De allele former af dette gen kan sĺ vćre identiske eller forskellige. Hvis de to allele former er identiske, er individet homozygot for den egenskab. Hvis de derimod er forskellige, er de heterozygote. Dvs:
Normalt, nĺr man skriver de allele former for et gen op, bruges som regel et enkelt bogstav for at gřre det lettere for os selv. Lad os antage, vi arbejder med et gen kaldet "a". Her skriver man den dominerende form med stort, dvs A og den recessive form med smĺt. Ved co-dom./inkomplet dom. skrives begge med stort, men man skriver noget andet i forbindelse med bogstavet, sĺ man kan skelne formerne. Dette vender jeg tilbage til senere. For at forstĺ, hvordan disse fire metoder fungerer, vil jeg introducere to ekstra begreber, nemlig genotype og fćnotype. Fćnotypen er det udseende, som individet har, mens genotypen er de allele former, som er til stede af genet, men som ikke nřdvendigvis kan ses via fćnotypen. Hvis vi tager et gen som eksempel, og kalder den ene allele form for A og den anden for a, sĺ kan der vćre fřlgende sammensćtninger (jeg bruger kun dominante og recessive allele former her (glem co-dom./inkomplet dom. et řjeblik): AA, Aa og aa
Ved co-dom./inkomplet dom. skriver man bare A+ og A1 f. eks, da de begge af lige dominerende. Co-dom./inkomplet dom. allele former for gener er meget udbredt ved f. eks kongepyton, Python regius. Hvis vi tager et gen som eksempel, og kalder den ene allele form for A+ og den anden for A1, sĺ kan der vćre fřlgende sammensćtninger (jeg bruger kun co-dom./inkomplet dom. allele former her): A+A+, A+A1 og A1A1.
Forskellen mellem co-dominant og inkomplet dominant er ikke sćrlig stor, men der er en tydelig signifikant forskel alligevel. Jeg har skitseret forskellen pĺ nedenstĺende figur. Ved A er der tale om inkomplet dominant og ved B er der tale om co-dominant. Forskellen er altsĺ, at ved inkomplet dominans bliver fćnotypen en sammenblanding mellem de to former og ved co-dominans bliver begge former udtrykt 100 %, men pĺ hver deres "del" af individet. Ved avl af udseende, som morfavl er, er der primćrt tale om inkomplet dominans. 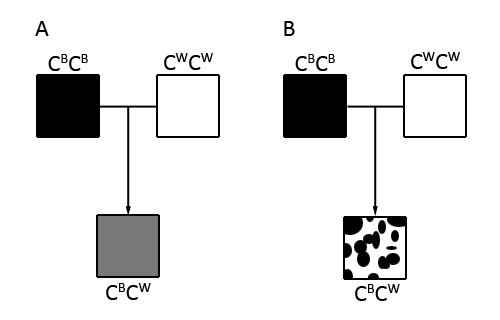
Figur 1: Inkomplet dominans eller co-dominans. A: Inkomplet dominans, B: co-dominans. HaplosufficiensHvad er det sĺ, der afgřrer, om et gen er dominant eller recessivt? Her skal vi tale om haplosufficiens. Der skal en bestemt mćngde genprodukt (typisk et protein) for at opretholde vildtypeudseendet. Pĺ figuren er denne grćnse symboliseret ved den rřde streg. Ved en heterozygot kan der ske to ting: 1, den ene velfungerende allel producerer nok genprodukt for at opretholde vildtypeudseendet, eller 2, den ene velfungerende allel producerer ikke nok genprodukt for at opretholde vildtypeudseendet. Situation af 1 er skitseret i den venstre del af figuren. Ved situation 1 er mutant-allelen derfor recessiv overfor vildtypen, da den ene velfungerede allel er nok til at se normal ud. Hvis der ikke produceres nok genprodukt, er mutantallelen enten dominant, inkomplet dominant eller co-dominant overfor vildtypen.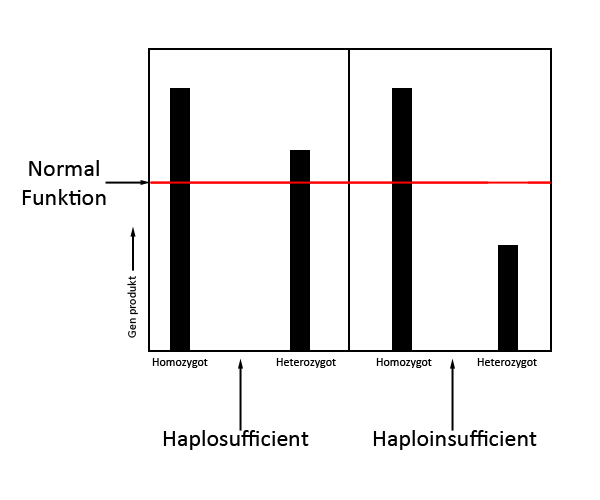
Figur 2: Haplosufficiens eller haploinsufficiens, forskellen mellem om en allel agerer recessivt, dominant, inkomplet dominant eller co-dominant overfor vildtypen. Nu skulle de vigtigste begreber vćre pĺ plads, og jeg vil gĺ ind i mere specifikke eksempler. Hvorfor er de to anery-former ikke kompatible? Hvad med de forskellige hypo-former?Jeg starter med de forskellige anery-former. Der er to forskellige
Bĺde anery og charcoal er reccesive i forhold til vildtypen, sĺ parres en anery med en charcoal, vil man fĺ unger med vildtypeudseende heterozygot for bĺde anery og charcoal. Nu vil jeg koncentrere mig om de forskellige hypo-former. Der er pĺ nuvćrende tidspunkt identificeret syv forskellige hypo-former. Jeg har inddelt disse syv former i fem forskellige grupper, nemlig A til E.
Note 1: Der er god evidens for at ultra-allelen oprindeligt stammer fra et hybridprojekt med Pantherophis spiloides. Nedarvning af generNu skal vi sĺ til at "beregne" afkom. Min foretrukne metode til dette er ved at bruge "Punnett Squares" (se figur 3), idet det giver et godt overblik over afkommet.Eksempel 1 – Et gen Vi starter med et simpelt tilfćlde, nemlig Normal het amel X Normal het amel. Systemet er lige til, man lister hannens alleler langs den ene akse, hvor jeg her arbitrćrt vćlger den vandrette akse, og hunnens langs den anden akse (dette vil jeg gřre i alle eksempler i denne tekst). Amel-locuset betegnes her som "A". 
Figur 3: Opbygning af et Punnett square med et enkelt gen. I de fire firkanter kan man sĺ se genotyperne pĺ afkommet, idet man kombinerer allelerne fra de to forskellige akser. Vi Fĺr AA, Aa, Aa og aa. Dvs. en amel og tre normalfarvede. To af disse normalfarvede vil vćre het amel, men da man ikke kan se pĺ udseendet, hvilke af de normale, der er det, fĺr de betegnelsen poss het amel 67 %. betegnelsen poss het, som stĺr for "possible heterozygot", dvs en mulig heterozygot. Hvis vi kigger i figur 3, kan vi se, at to ud af de tre individer med fćnotypen A er heterozygot. Dette betegnes "possible het 67 %" fordi der er 2/3-dele chance for den er het. Der er kun to muligheder for procentangivelser ved possible het, nemlig 50 % og 67 % (som nogle gange ses betegnet som 66 %, men hvis man kender afrundingsreglerne i matematik er 67 % mere korrekt, hvis man ikke vil bruge brřken 2/3). Eksempel 2 – To gener I dette her eksempel tilfřjer vi et ekstra gen. Her tager vi eksemplet Normal het amel, anery X Normal het amel, anery. anery-locuset betegnes her som "B" Nĺr man arbejder med to eller flere gener kan man sćtte sine Punnet Squares op pĺ to forskellige mĺder, som pĺ figuren hedder henholdsvis A og B. Ved metode A gřr man som i eksemplet ovenover, dvs lister hannens og hunnens alleler langs de to akser. Ved metode B lister man afkommet fra hvert gen langs hver akse. Jeg foretrćkker personligt metode A, hvorfor jeg vil bruge den metode i de senere eksempler, men B metoden giver det samme resultat, sĺ den er lige sĺ rigtig at bruge. 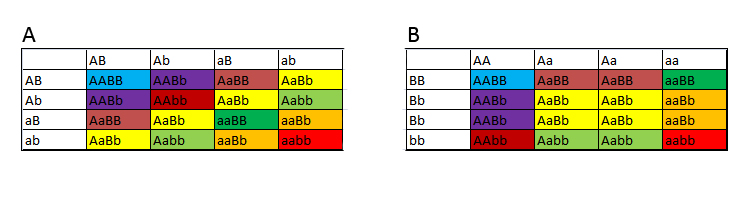
Figur 4: To muligheder for at opbygge et Punnett square med to forskellige gener. Igen kan vi hurtigt aflćse de forskellige geno- og fćnotyper ud fra skemaet. Jeg vil dog lave et ekstra eksempel. Eksempel 3 – Tre eller flere gener Dette tilfćlde tager jeg primćrt med for at vise, hvor vanskeligt det er at arbejde med mere end to gener. Ved to gener er sandsynligheden for at fĺ det řnskede resultat 1/16, hvilket er ganske pćnt - med 3 eller flere gener bliver det langt vćrre. Jeg bruger eksemplet Normal het amel, anery, stripe X Normal het amel, anery, stripe. Her kalder vi stripe-lucuset for "T". 
Figur 5: Et Punnett square som viser hvor lille sandsynlighed der er for det řnskede resultat ved tre gener. Som man kan se, er det kun 1/64, som bliver det řnskede resultat (som er aabbtt, markeret med rřdt). Hvis man arbejder med fire gener er sandsynligheden helt nede pĺ 1/256 (se figur 6). Dette er ogsĺ grunden til, at der er mange kombinationsmorfer, som bestĺr af to forskellige morfer. Kombinationsmorfer, som bestĺr af tre forskellige morfer, er ogsĺ ved at blive fremavlet i střrre mćngder, mens firedobbelte morfer stadig er forholdsvis sjćldne (de bliver dog lavet). Hvis man vil arbejde med 3-4-5 dobbelte kombinationsmorfer, kan man gĺ ud fra individer som er homozygot for 1-2 forskellige morfer og heterozygot for de sidste, da man derved har en hřjere sandsynlighed for at opnĺ det řnskede resultat. 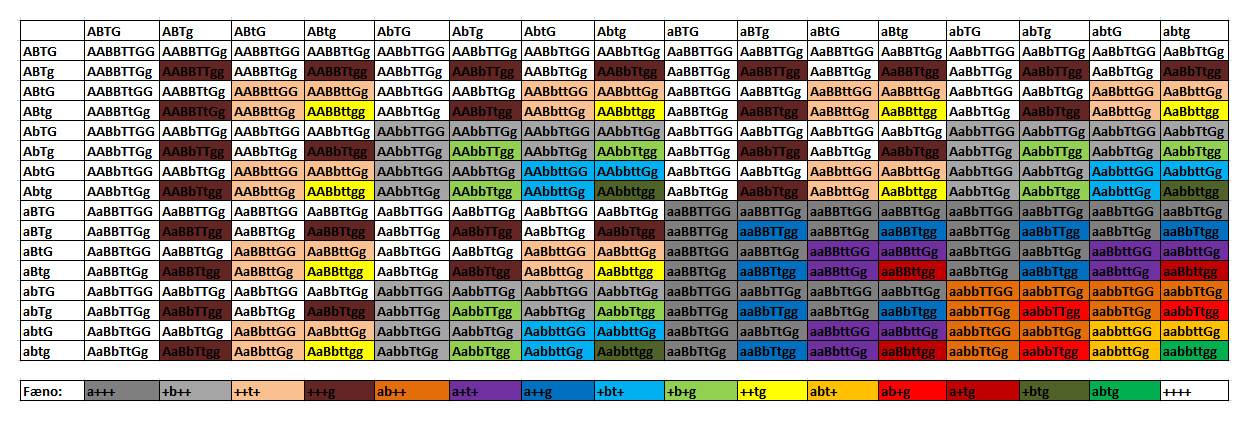
Figur 6: Et Punnett square som viser hvor lille sandsynlighed der er for det řnskede resultat (aabbttgg) ved fire gener. Dog kan man ogsĺ se at der vil komme mange forskellige individer ud, hvilket samtidig kan gřre det til et interessant projekt. Eksempel 4a - Dominante gener Střrstedelen af morferne ved kornsnoge er bestemt af gener hvor mutanten er reccesiv til vildtypen. Der er dog fundet (indtil nu) to gener hvor mutanten er dominerende overfor vildtypen. Disse er ved skrivningen af denne tekst dog endnu ikke fundet ved til Danmark, i hvert fald ikke hvad jeg er vidende. De vil dog komme pĺ et tidspunkt, sĺ beskrivelsen kan lige sĺ godt komme online nu. Jeg tager her eksemplerne Tessera het vildtype X Tessera het vildtype (A), Tessera het vildtype X Tessera (B), Tessera het vildtype X Vildtype (C), Tessera X vildtype (D) og Tessera X Tessera (E). Tesserlocuset kalder jeg her for D. I dette tilfćlde vil vidtypen hedde d+ mens mutantallelen vil hedde DD 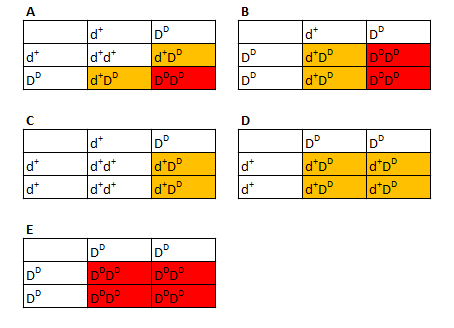
Figur 7: Eksempler pĺ Punnett squares over muligheder af afkom ved et enkelt dominant gen. I figuren har jeg farvet Tessera het vildtype for orange og Tessera (homozygot) for rřd. Da dette er et dominant gen kan man dog ikke se forskel mellem disse, sĺ reelt burde man betegne dem som poss het. I figur 7(A):Tessera poss het vildtype (67 %), figur 7(B):Tessera poss het vildtype (50 %) . I figur 7(C) og 7(D) ved man med sikkerhed at der er tale om Tessera het vildtype Eksempel 4b - Dominante gener sammen med recessive gener Dette eksempel burde ikke vćre sĺ svćrt selv at beregne, jeg tager det dog med alligevel for at vise et eksempel med bĺde recessive og dominante gener. Jeg tager eksemplet Tessera het vildtype, amel, anery X Ameltessera het vildtype. For at gřre skemaet lettere lćseligt vil jeg ved amel og anery locuset blot bruge A/a og B/b som tidligere og +/D ved Tessera. Dvs. symbolerne for vildtypeallelen er henholdsvis A, B og + mens mutantallelen er a, b og D. 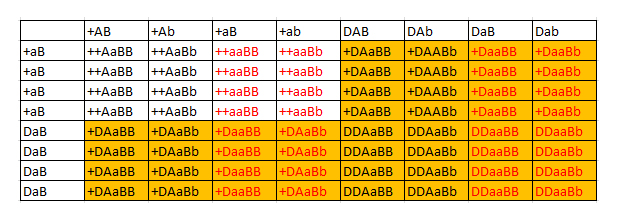
Figur 8: Et Punnett square som viser hvordan man regner med dominerende alleler sammen med recessive alleler. I figuren er individer der fćnotypisk er Tessera farvet orange, mens de individer der er amel er teksten farvet rřd. Som man kan se kommer der ikke nogle anerys, hvilket er fordi kun den ene forćlder bćrer dette gen. I dette eksempel fĺr vi i alt fire forskellige fćnotyper, nemlig vildtype, amel, tessera og ameltessera. Vi ved dog mere om dem end det. Ud fra forćldrenes genotyper ved vi at alle de individer der ikke er amel er het amel. Ligeledes mĺ alle ungerne vćre poss het anery (50 %). Tesseraindividerne mĺ vćre poss het vildtype (67 %). Vi kan altsĺ skrive dem op som: Vildtype het amel poss het anery (50 %) Amel poss het anery (50 %) Tessera het amel, poss het vildtype (67 %) anery (50 %)(note 2) Ameltessera poss het vildtype (67 %), anery (50 %)(note 2) Note 2: Disse dominante gener ved kornsnoge er meget nye, sĺ der er ikke fundet en entydig mĺde at navngive dem pĺ endnu. En mulighed er at gřre som jeg har skrevet ovenfor, men det betyder ikke at det vil bliove praksis. En mulighed ville vćre at kalde en Tessera het vildtype for "Tessera" og en homozygot tessera for "Homo tessera" - dvs lidt det samme princip betegnelsen "super" ved mange kvćlere, men dog stadig med en betegnelse som giver mening set med en genetikers řjne. Ved at bruge denne model ville man ved poss het kunne kalde dem "Poss Homo tessera". Hvordan navngivningen kommer til at ske i praksis er dog tidligt at spĺ om pĺ nuvćrende tidspunkt, sĺ dette er blot nogle af mine tanker omkring emnet. Eksempel 5 - Flere forskellige alleler af samme gen Dette er et emne, som er meget relevant, og med den sandsynlige introduktion af forskellige hypo-alleler i Danmark indenfor en ĺrrćkke, et emne som bliver mere vigtigt at forstĺ. Der er to loci som er relevante her, nemlig "motley" som har allelerne "vildtype, motley og stripe". Den anden er "hypo" med allelerne "vildtype, christmas, hypo og strawberry" (Den er dog ogsĺ relevant ved amellocuset - situationen er dog helt identisk med hypolocuset, men pga. tvivlen omkring ultraallelens ophav vil jeg ikke gĺ ind i denne situation her). Ved motley locuset er vildtypen dominerende overfor begge mutantalleler, men motleyallelen er dominerende overfor stripeallelen. Dvs. man kan reelt skrive allernes dominans op som: vildtype>motley>stripe. Ved hypo locuset er situationen en anden, vildtypen er stadig dominerende overfor alle tre mutantalleler, men mutantallelerne er indbyrdes inkomplet dominant overfor hinanden, dvs.: vildtype>christmas=hypo=strawberry. Jeg vil fřrst tage et par eksempler med motleyallelen, nćrmere bestemt Motley het stripe X Motley het stripe(note 3) (A) og Motley het stripe X Vildtype het stripe (B). Jeg kalder her motley locuset for M. 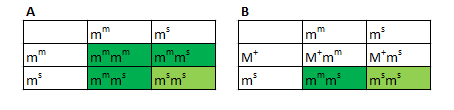
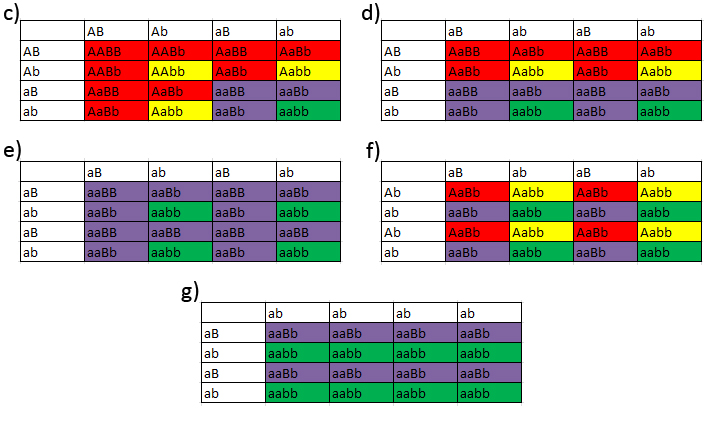
Figur 9: Eksempler pĺ Punnett squares ved et gen med mere end to forskellige allele former. I figuren har jeg farvet individer der er fćnotypisk motley mřrkegrřn, mens fćnotypisk stripe er farvet lysegrřn. I (A) vil man blive nřdt til at betegne motleysne som Motley poss het stripe (67 %) og i (B) som Motley poss het stripe (50 %). I B kommer vi ud for en ny situation med de normale. Vi kan se de normale er heterozygoter, men ikke hvilken allel de har sammen med vildtypen. Dette klarer man ved at betegne dem som Vildtype het motley eller stripe Nu vil jeg sĺ tage nogle eksempler med de forskellige alleler pĺ hypolocuset. Jeg vil tage eksemplerne Vildtype het hypo X Vildtype het strawberry (A), Hypo X Vildtype het strawberry (B), Hypostrawberry X Hypochristmas (C) og Hypostrawberry X Vildtype het christmas (D). 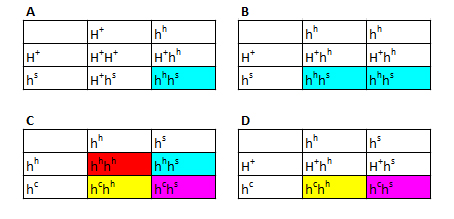
Figur 10: Eksempler pĺ Punnett squares ved et gen med mere end to forskellige allele former. Farvemćssigt i figuren har jeg valgt at benytte de additive primćrfarver til de tre alleler, sĺ christmas=rřd, hypo=grřn og strawberry=blĺ. Ved heterozygoterne har jeg valgt at fřlge dette farvesystem, sĺ christmas+hypo=gul, christmas+strawberry=magenta og hypo+strawberry=cyan. Som man kan se er der mange muligheder ved et sĺdan system. Fordelen her fremfor ved motleylocuset er at man kan identificere heterozygoterne visuelt, bortset hvis en af allelerne er vildtypen, dvs., man kan se pĺ dyret om der er tale om en homozygot hypo, eller en hjypostrawberry eksempelvis. Man fĺr altsĺ kun brug for begrebet "poss het" ved normaltudseende dyr. En anden ting vćrd at notere er at man kan fĺ mange forskellige fćnotyper ud, eksemplet (C) viser det glimrende. Forćldredyrene har vćrd deres fćnotype, som man ogsĺ fĺr ved afkomene, men samtidig fĺr man to ekstra fćnotyper, og alt dette sker ved blot ét gen! Note 3: Oftest ser man et individ med en motleyallel og en stripeallel betegnet som "motley het stripe". Dette betyder blot at individet har fćnotypen motley, men at den bćrer de to forskellige mutantalleler pĺ motleylocuset. Man vil se jeg nogen gange har skrevet vildtype, andre gange normal. Dette er to begreber som begge er lige rigtige. Grunden til skift imellem dem er jeg har skrevet teksterne pĺ forskellige tidspunkter, og nogen dage har jeg skrevet vildtypen og andre dage normalen. Gensymboler ved kornsnogeI denne liste nćvner jeg brugte gensymboler ved kornsnoge. Alle disse er kopieret fra bogen Cornsnake Morph Guide - 2011 versionen skrevet af Charles Pritzel. Der er ikke blevet udgivet en bog i 2012, sĺ de nyeste avlsresultater vedrřrende palmetto, pied-sided og tessera har derfor ikke vćret i betragtning til denne liste.
Note 4: Pĺ tidspunktet denne liste blev beskrevet havde man endnu ikke reproduceret denne morf. I 2011 blev den bevist til at vćre arvelig, og mutantallelen til at vćre recessiv til vildtypen. Note 5: Pĺ nuvćrende tidspunkt er genetikken ved Pied-sided genet ikke forstĺet fuldstćndigt. Man ved der er tale om et gen hvor mutanten er recessiv til vildtypen. Det man endnu ikke har kortlagt er om det er en ny allel af diffusion-genet eller om det er en nyt gen, som kun kommer fćnotypisk til udtryk sammen med Diffusion. Yderligere avlsresultater forsřger at kortlćgge dette. Note 6: Pĺ tidspunktet denne liste blev beskrevet vidste man ikke om mutantallelen var dominerende, eller inkomplet/co-dominerende til vildtypen. Denne er dog efterfřlgende blevet vist til at vćre dominant. Typiske fćno-type fordelingerVed avl af genetiske trćk vil man efterhĺnden se, at der er et mřnster over forholdene mellem de nedarvede gener. Pga, sĺ mange muligheder for resultater ved mange gener, vil jeg kun fokusere pĺ 1 og 2 gener.Et gen:
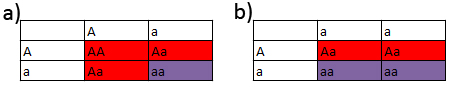
Figur 11: Fćnotypefordelinger ved 1 gen. a: Her ses tre individer med en fćnotype og et individ med en anden fćnotype, dvs en 3:1 fordeling. Samme princip er tilfćldet ved illustrationerne 7b og 8c-g. To gener:
Figur 12: Fćnotypefordelinger ved to gener. ModifikationsgenerEt modifikationsgen er, som navnet antyder, et gen, som modificerer et anden gen. Dette modificeringsgen har ingen anderledes fćnotype i sig selv, kun sammen med et andet gen.I figuren nedenunder betragter vi to gener, rřd og blĺ. I A er der blot tale om to forskellige reccesive gener, og vi ser den forventede 9:3:3:1 fćnotype-fordeling. I B er situationen lidt anderledes. Det er nřjagtig samme parring, men blĺ er et modifikationsgen af rřd. Dette betyder at blĺ kun kommer til udtryk nĺr rřd ogsĺ er ved ydtryk. Her fĺr vi i stedet en modificeret 9:3:3:1 fordeling, nemlig 12:3:1. Genotyperne i A og B er 100 % identiske. Ved modifikationsgener er det udelukkende fćnotypefordelingen der bliver ćndret 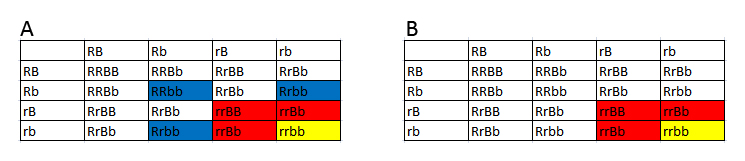
Figur 13: To Punnet squares som viser forskellen mellem to normale gener og et normalt gen plus et modifikationsgen. Man har mistanke om et enkelt modifikationsgen ved kornsnoge, nemlig pied-sided. Teorien er her, at pied-sided er et modifikationsgen af diffused, dvs. at man kun kan fĺ pied-sided udtrykt samtidig med diffused. Man har endnu ikke lavet nok testparringer til at verificere eller falsificere denne mistanke, men der bliver arbejdet meget pĺ denne front. Křnsbunden nedarvningInden jeg gĺr dybt ned i dette emne, er der nogle informationer, man skal huske pĺ. Der er flere forskellige systemer som hanner og hunner adskilles pĺ i dyreverdenen. Jeg nćvner her tre forskellige systemer. I listen nedenunder har jeg nćvnt hannerne fřrst og hunnerne sidst.
Jeg vil herefter kun betragte ZW-systemet, da det er det eneste relevante ved kornsnogeavl. Eksempel 1 - Z-bundet reccesivt gen Til at starte med betragter vi et gen, som er bundet til Z-kromosomet, jeg kalder den her Zr. Lad os antage, at denne allel giver dyret en rřd farve. Vildtype-allelen kalder jeg Z+. Zr agerer reccesivt til vildtypen. Ved hannerne fungerer den som de gener, vi ellers kender. Som heterozygot kommer den ikke til udtryk, og den kommer til udtryk som homozygot. Ved hunner er det lidt anderledes. Hunner har kun et Z-kromosom, sĺ hvis den fĺr denne allel, sĺ kommer det til udtryk idet hunnen er hemizygot for dette trćk. I figuren har jeg vist alle de mulige kombinationer der findes. I figuren reprćsenterer en hvid firkant normalt udseende, en orange firkant normalt udseende, men som er heterozygot, og en rřd firkant som et individ som udtrykker trćkket. Der findes ingen kendte eksempler pĺ křnsbundne gener ved kornsnoge, som er relevante for morfavl. Hos mennesket er blřdersygdommen et eksempel pĺ et křnsbundet gen. 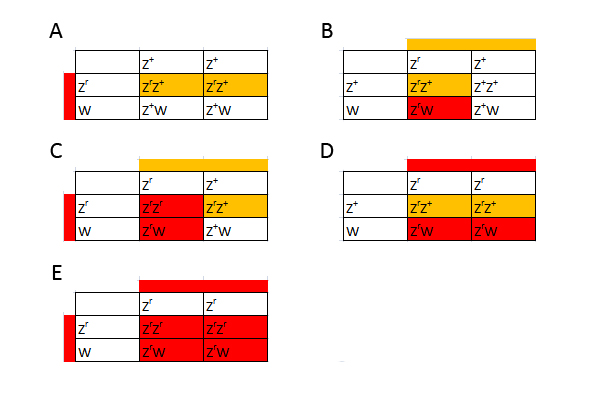
Figur 14: Punnett squares som viser muligheden for et Z-koblet reccesivt gen. Som man kan se ud af figuren, kan man konkludere nogle bestemte ting. Ud fra hunnerne kan man konkludere fřlgende: Hvis alle hunnerne i et kuld bliver rřde, ved vi at hannen er homozygot for trćkket. Hvis halvdelen af hunnerne bliver rřde, ved vi at hannen er heterozygot for trćkket. Ud fra hannerne kan man konkludere fřlgende: Hvis alle hannerne i et kuld bliver rřde ved vi, at hannen er homozygot for trćkket, og hunnen er hemizygot for trćkket. Hvis halvdelen af hannerne bliver rřde, ved vi, at hannen er heterozygot for trćkket, og hunnen er hemizygot for trćkket. Eksempel 2 - Z-bundet dominant gen I det nćste eksempel er det igen et Z-bundet gen men hvor jeg antager allelen er dominant til vildtypen. Jeg kalder det ZB, som giver en blĺ farve. I figuren vil de hvide firkanter igen reprćsentere normalt udseende. Begge blĺ farver er et individ som udtrykker trćkket, de lyseblĺ er blot heterozygote 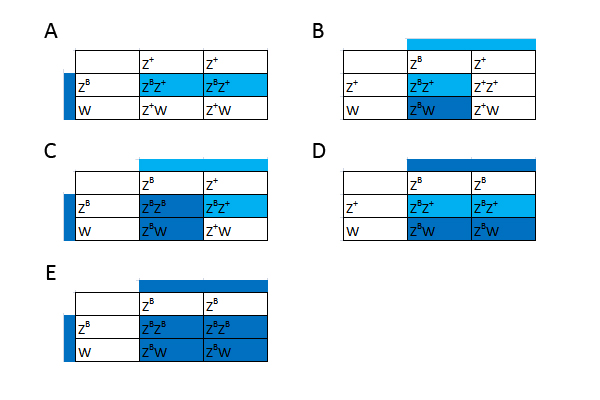
Figur 15: Punnett squares som viser muligheden for et Z-koblet dominant gen. Man kan igen konkludere nogle ting ud fra figuren. Ud fra hunnerne kan man konkludere fřlgende: Hvis alle hunnerne i et kuld bliver blĺ ,ved vi, at hannen er homozygot for trćkket. Hvis halvdelen af hunnerne bliver blĺ, ved vi, at hannen er heterozygot for trćkket. Hannerne kan man ikke sige lige sĺ meget til. Dog kan vi sige, at hvis halvdelen af hannerne bliver blĺ, mĺ hannen vćre heterozygot. Som nogle mĺske har lagt mćrke til, har jeg ikke givet noget eksempel hvor trćkket er inkomplet dominant. Nedarvningen er det samme som i de andre to eksempler. Hannerne har dog tre forskellige fćnotyper, mens hunnerne kun har to (hvis vi antager der ikke er "Z-silencing" ligesom man ved andre dyr ser "X-silencing" ved XY-systemet), idet de ikke kan vćre homozygot for allelen. Hvilken mĺde, hunnerne ser ud pĺ, kan vćre svćr at forudsige, idet der kun er en enkelt kopi til stede af et gen i stedet for to. Jeg gennemgĺr altsĺ ikke et tćnkt eksempel, idet der er mange variable, man skal tage hřjde for, og som ikke nřdvendigvis kan besvares, fřr man har et reelt eksempel at studere – hvilket ikke findes ved kornsnoge. Hvis vi stod med et W-bundet gen er det en meget nem sitation. Hvis hunnen har mutationen, vil hun give den videre til alle hunner. Hanner vil ikke kunne arve denne mutation. Koblet nedarvningKoblet nedarvning er lidt mere kompliceret, men jeg vil prřve at forklare det alligevel. Under meiosis, som er en specifik form for celledeling, som er nřdvendig for sexuel reproduktion, er der et tidspunkt, som kaldes "fire-kromatid tidspunktet". Ved dette tidspunkt er begge kromosomer blevet duplikeret og dannet to par sřster kromatider. Disse to sřsterkromatider er placeret overfor hinanden, inden de fordeles til hver deres celle. Hvis der sker et overkryds mellem sřsterkromatiderne (dette kaldes homolog rekombination), bliver to af kromatiderne en kombination af den ene og den anden. I figur 10 har jeg skitseret situationen. Her er det vigtigt at huske pĺ, at denne proces sker i den ene forćlder, da produktet af meioses bliver 4 haploide celler (haploid betyder celler som kun indeholder et enkelt kromosomsćt, hvorimod diploid indeholder to kromosomsćt). Den meiotiske celledeling foregĺr i begge forćldre, og ved reproduktion bliver en haploid celle fra faderen fusioneret med en haploid celle fra moderen. Dette danner derfor en diploid celle, som vil vokse op til et nyt individ.I figuren har jeg farvet det ene kromosom rřdt og det andet blĺt for at gřre det lettere at overskue situationen. De to alleler fra hvert kromosom vi er interesserede i er farvet sort eller hvidt. 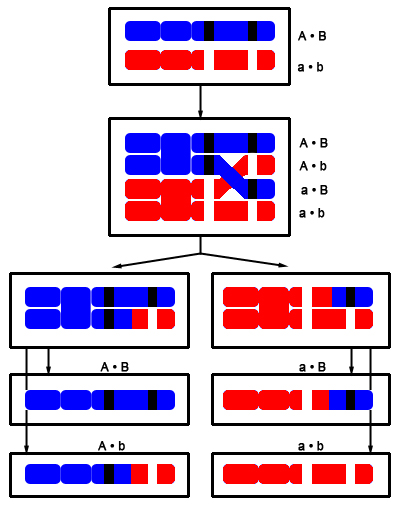
Figur 16: Koblet nedarvning Her kan man se, at forćldretyperne hedder a•b og A•B, mens mulighederne A•b og a•B er rekombinante, som kun forekommer nĺr der er foretaget et overkryds ("•" betyder de er koblet til hinanden). Sandsynligheden for et sĺdan overkryds afhćnger af afstanden mellem de to loci, jo tćttere pĺ hinanden jo mindre sandsynlighed. For at tage et eksempel kan vi antage, at A og B lociene er koblet til hinanden, og den ene forćlder har genotypen a•b,a•b, mens den anden har A•B,a•b. Den normale har de to mutant alleler pĺ det samme kromosom, dvs A•B og a•b. Hvis vi parrer disse to sammen, og der ikke sker overkryds, vil vi kun se to fćnotyper; ab og AB. Ved overkryds kan vi derimod ogsĺ fĺ fćnotyperne aB og Ab frem. Eksempel 1 (ingen overkryds): Forćlder et hedder a•b,a•b, dvs den kan kun give a•b videre til sit afkom. Forćlder to hedder A•B,a•b, dvs den kan give enten A•B eller a•b videre (idet der ikke sker overkryds). Afkommet kommer derfor til at hedde enten A•B,a•b eller a•b,a•b, hvilket betyder vi kun ser to fćnotyper, vildtypen (AB) eller dobbeltmutanten (ab) Eksempel 2 (mulighed for overkryds): Forćlderne hedder det samme som i eksempel 1. Dvs forćlder et kan stadig kun give a•b videre (idet den er homozygot for begge gener). Forćlder to kan, som i fřrste eksempel, give A•B og a•b videre hvis der ikke sker overkryds, hvis der derimod sker overkryds, kan den ogsĺ give kombinationerne A•b og a•B videre, hvorfor vi her kan se flere fćnotyper, nemlig AB, ab, Ab og aB. Selektiv avlSelektiv avl adskiller sig fra den type avl, der ellers er blevet diskuteret pĺ et vćsentligt punkt; den fřlger ikke-genetisk-nedarvning (som alle tidligere emner har fulgt). Dette er fordi der ikke er tale om et enkelt gen, men en kombination af mange forskellige. Det man gřr i stedet er at forćdle linjen i flere generationers avl.Hvis man f. eks gĺr efter at lave en candy cane, som er en rřd/hvid kornsnog, og man har almindelige amelanistiske som udgangspunkt tager man de unger, som udviser mest af det trćk, man vil have, og parrer sammen - ud fra deres unger gřr man det samme, og dette fortsćtter man i generationer, indtil det řnskede resultat. Situationen er skitseret i figur 17. 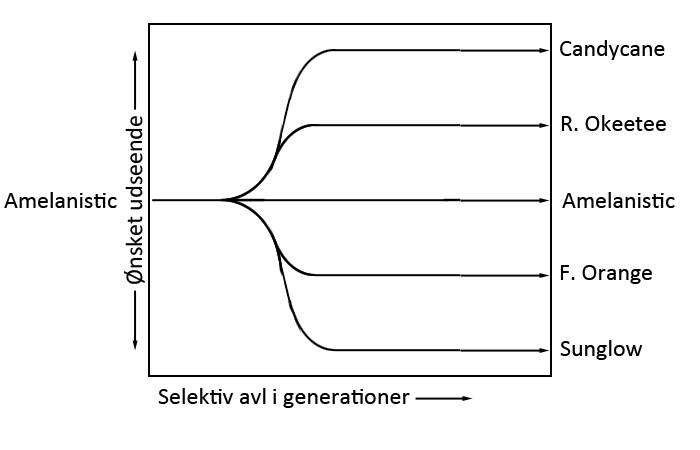
Figur 17: Selektiv avl. Morfer i naturenSelvom mange af morferne er indfanget i naturen (se nćste sektion) betyder det ikke, at de normalt trives der. Eksempelvis kan mangel pĺ camouflage gřre et individ mere sĺrbart overfor naturlige fjender. Der lever dog en naturlig population af anerytristiske kornsnoge i det sydvestlige Florida. Det samme er tilfćldet i Georgia. Dette tyder derfor pĺ, at netop den anerytristiske form ikke er mere sĺrbar, end vildtypen er.Hvor fĺr man morferne fra?Ja, dette er et interessant spřrgsmĺl. Mange af dem er reelt mutationer som er sket spontant i et individ i naturen, som herefter er blevet indfanget og avlet videre pĺ. Mutationer, som med sikkerhed er sket i naturen, er bl.a. amel, anery, charcoal, caramel, lava, palmetto, sunkissed, terazzo... Noget ekstra interessant er, at det charcoalindivid, som blev indfanget i naturen, faktisk var heterozygot anery. Dette ville man normalt betegne som meget usandsynligt, men netop anery har vist sig ikke at vćre en defekt for dyrene i naturen, i og med at der lever en population af anery kornsnoge. Charcoal individet blev ogsĺ indfanget i dette omrĺde.Nogle gener er dog ogsĺ dukket op under avl i fangenskab. Dette er f.eks tilfćldet med lavender AfslutningHvis du har yderligere eller opklarende spřrgsmĺl omkring emnet, er du velkommen til at kontakte mig (Thomas) pĺ mail thomas@g-reptiles.com eller thomas@grochalski.dkDenne tekst er beskyttet af ophavsretten og mĺ ikke benyttes andetsteds uden tilladelse. © Thomas Grochalski 2005-2012. No part of this website may be reproduced elsewhere without our express written permission. Website designed by Thomas Grochalski |